Kde se tvoří spory v rostlinách?
§ 32. Střídání způsobů rozmnožování a generací v životním cyklu rostlin
*Tvorba zárodečných buněk a oplodnění u krytosemenných rostlin
V § 29-3 jste se seznámili s nepohlavním rozmnožováním rostlin a nyní se budeme podrobněji zabývat pohlavním rozmnožováním rostlin na příkladu krytosemenných rostlin.
Strukturu a funkce generativního orgánu krytosemenných rostlin – květu, již znáte. Podívejme se na procesy probíhající v prašníku tyčinky a vajíčku pestíkového vaječníku.
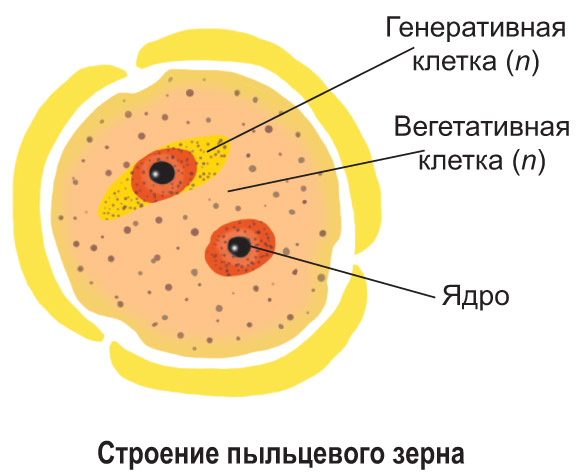
Prašník obsahuje pylová hnízda (sporangia), ve kterých se tvoří výtrusy. Z každé mateřské buňky se dělením vytvoří čtyři jednobuněčné haploidní spory. Poté je každá spora rozdělena na dvě buňky: velkou vegetativní a malou generativní. Jsou pokryty dvojitou hustou skořápkou s póry a tvoří se pylové zrno (samčí pohlavní generace). Po rozdělení generativní buňky se vytvoří dvě spermie (mužské reprodukční buňky bez bičíků). Všechny buňky v pylovém zrnu jsou haploidní.
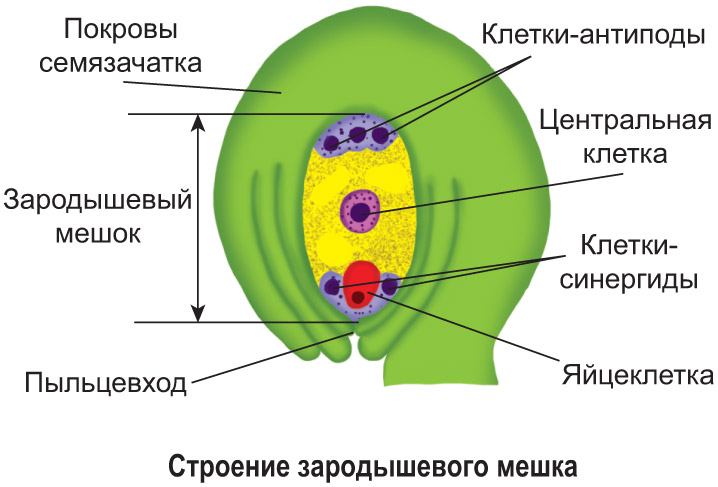
Část květu, která určuje ženské pohlaví, je palička . Skládá se z stigma, styl и vaječníků (viz obrázek v § 0-4). Vaječník obsahuje vajíčko (vajíčko), které má na vnější straně integument. Na vrcholu vajíčka se stélka nesrůstá a vytváří se pylový průchod. Jedna z mateřských buněk vajíčka v blízkosti pylového otvoru se zvětšuje, dělí a tvoří čtyři haploidní spory. Tři z nich zemřou. Ze čtvrté spóry se po několika děleních jádra a cytoplazmy vytvoří sedm buněk. Na každém pólu jsou umístěny tři buňky s haploidní sadou chromozomů a mezi nimi je velká centrální diploidní buňka. Jedna ze tří buněk na pólu poblíž pylového kanálku se stává vaječnou buňkou. Dvě sousední buňky se nazývají synergické buňky a tři buňky na opačném pólu se nazývají antipodní buňky. Výsledná struktura sedmi buněk s vajíčkem představuje zárodečný vak (ženské pokolení).
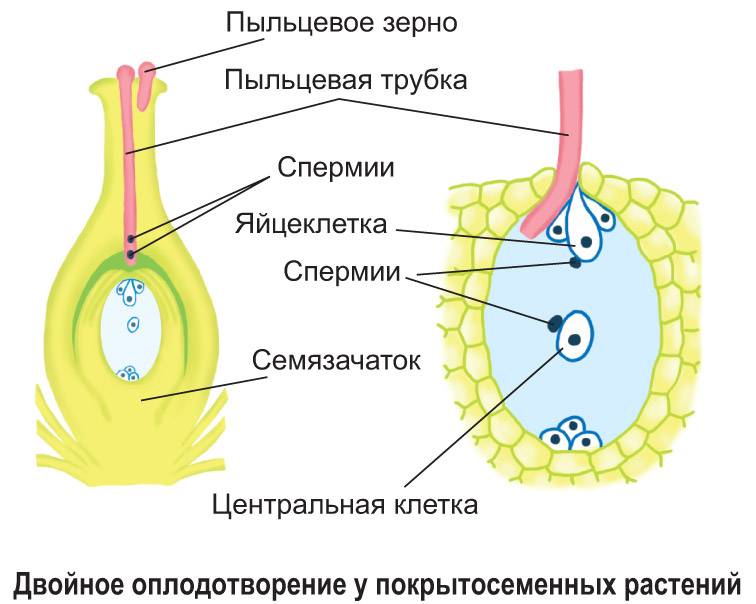
Přenos pylových zrn z prašníku tyčinky na bliznu pestíku u krytosemenných rostlin je tzv. opylení. Lze ji provádět pomocí hmyzu, větru, vody, ptáků nebo samoopylení. Po dosednutí na bliznu pestíku pylové zrno vyklíčí: vegetativní buňka se rozšíří do dlouhé pylové láčky, která rozpustí tkáň stylu, pronikne do dutiny vaječníku a pylovým kanálkem se dostane do zárodečného vaku. Spermie se pohybují podél pylové láčky a vstupují do embryonálního vaku. Jeden z nich oplodní vajíčko zárodečného vaku a druhý splyne s centrální buňkou.
Tento typ oplodnění objevil v roce 1898 ruský cytolog S. G. Navashin a byl nazýván „dvojitým oplodněním“. Posloupnost jeho fází je uvedena v diagramu.
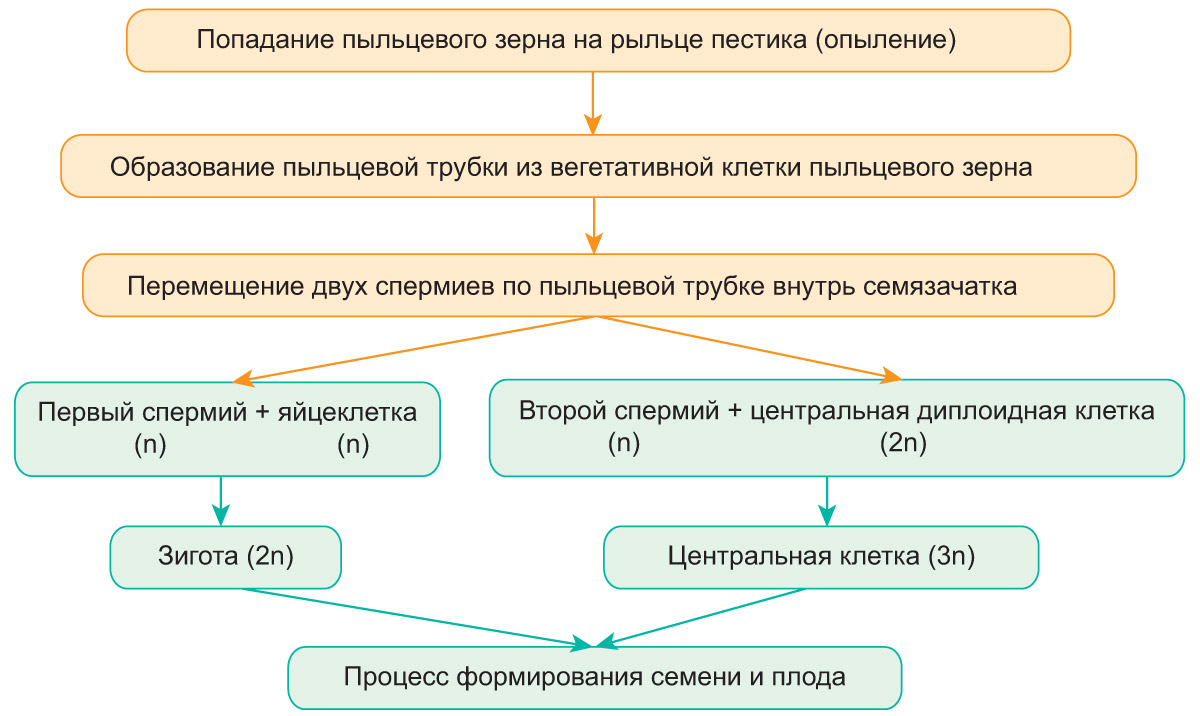
Po vytvoření zygoty synergidy odumírají v zárodečném vaku. Centrální triploidní buňka se opakovaně dělí, což má za následek tvorbu endospermu, který obsahuje zásobu živin. Embryo semena se tvoří ze zygoty. Po jeho vzniku antipody umírají. Tvorba semene a plodu je znázorněna na obrázku.
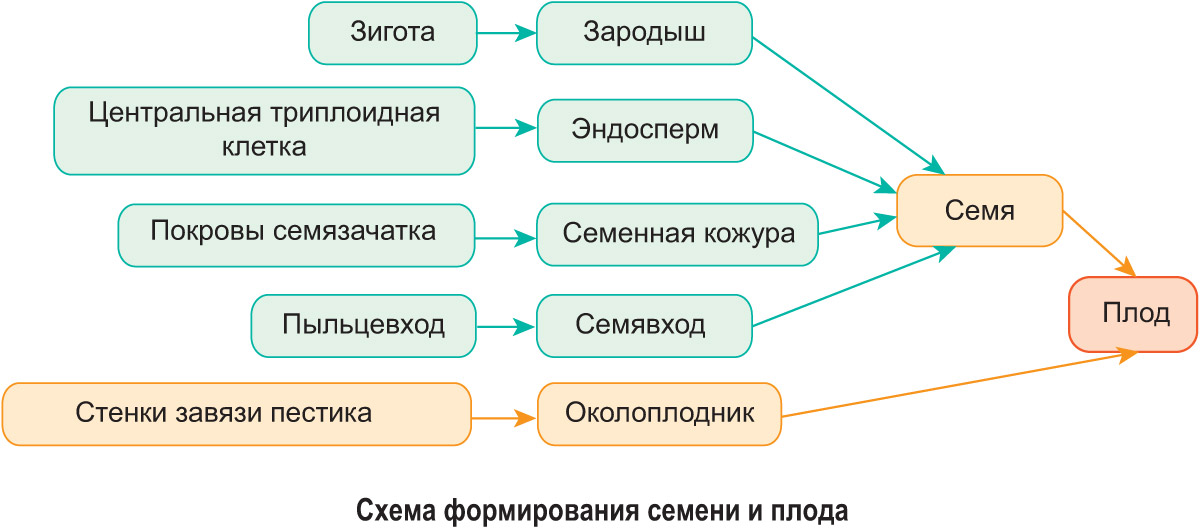
U některých rostlin se na tvorbě oplodí může podílet kalich a schránka.
Z vajíčka se tedy tvoří semeno a plod se vyvíjí na místě vaječníku.
Biologický význam dvojitého oplodnění je velmi velký. Na rozdíl od nahosemenných, kde se endosperm vyvíjí před oplodněním, u krytosemenných rostlin se endosperm tvoří až po oplození. To přináší významné úspory energetických zdrojů. Endospermové buňky obsahují triploidní sadu chromozomů, což vede ke zvětšení velikosti buněk a množství živin, které zvyšují odolnost embrya vůči nepříznivým faktorům.
Výtrus je obecný název pro rozmnožovací orgány hub a nižších rostlin. Skládá se z jedné nebo více buněk, které jsou odděleny od těla matky a jsou určeny k reprodukci. Zároveň se používají další názvy, které charakterizují účel, původ nebo jiné charakteristické vlastnosti [4].

Spory hub a houbám podobných organismů
Spory hub a houbám podobných organismů

Houby a houbám podobné organismy jsou schopny produkovat obrovské množství spor, jejichž počet může dosahovat až několika set milionů z několika centimetrů kolonie. Obrovských rozměrů však dosahuje i úbytek reprodukčních prvků, neboť pouze jednotlivé spory jednoho exempláře stihnou vyklíčit a zplodit potomstvo [3] .

Spóry plísní třídy Classa
Basidiomycetes (Basidiomycety)
Spóry plísní třídy Classa
Basidiomycetes (Basidiomycety)

2. Basidiospora Insolibasidium deformans
Typy spór hub a houbám podobných organismů
Spory plísní jsou obvykle nejméně variabilní struktury. Ve tvaru, velikosti a dalších charakteristikách jsou pro každý druh relativně konstantní a jsou důležitou charakteristikou pro identifikaci a klasifikaci hub a houbám podobných organismů [3].
Spory plísní se podle vzhledu a struktury dělí na dvě velké skupiny – pohyblivé a nepohyblivé [3].
Pohyblivé jsou zoospory. Mají vzhled nahých hrudek protoplazmy, bez skořápky, vybavené bičíky (motorickým aparátem). Zoospory se tvoří především v organismech úzce spjatých s vodním prostředím a jsou dobře přizpůsobeny pohybu v něm [1] [3].
Fixní výtrusy mají skořápku a postrádají motorický aparát. Pohyblivé výtrusy jsou velmi rozmanité ve tvaru a struktuře. Každá nepohyblivá spora však začíná jako jednobuněčný útvar. Je to jednoduchá buňka, ve většině případů kulovitého, vejčitého nebo elipsoidního tvaru. Často, jak spor roste, zvětšuje se, ale zachovává si tvar získaný v době svého výskytu. V ostatních případech se tvar spory může změnit [3].
Hlavní typy nepohyblivých spor:
- Zygospora je sexuální spóra hub třídy Zygomycetes.(Zygomycetes), vzniklý při procesu zygogamie, vyklíčí po období klidu ve speciální embryonální sporangium [5] .
- Sporangiospora je endogenní nepohyblivá spora, pokrytá schránkou a vytvořená uvnitř sporangia, která se vyvíjí na specializované hyfě (sporangioforu), která obvykle vystupuje nad substrát [5]. Sporagiespooras provádějí nepohlavní rozmnožování druhů třídy Zygomycetes(Zygomycetes)[5].
- Konidie jsou exogenní výtrusy nepohlavního rozmnožování, vznikající na povrchu výtrusných orgánů (na konidioforech) [3] ;
- Chlamydiospora je silnostěnná buňka, modifikace mycelia, schopná dát vznik novému myceliu [5].
- Cysta je klidová spora vytvořená pohlavně, s hustou, někdy vícevrstevnou schránkou [4];
- Oospore je sexuální stádium oogamie, charakteristické pro druhy třídy Oomycetes (Oomycetes), ve zralém stavu má dvouvrstvý obal, obsahuje ooplast a velké množství lipidových látek v cytoplazmě [2];
- Askospora je endogenní, haploidní spora vytvořená uvnitř ascus po mitotickém dělení postmeiotických jader [5].
- Teliospora (ustospora, sněťová spora) je silnostěnná klidová buňka, která tvoří teliobasidium, charakteristické pro druhy třídy Urediniomycetes(Urediniomycetes)[5].
- Spermace je forma sporulace hub z řádu Rustaceae (Uredinales), předcházející vzniku aeciospor [5].
- Aeciospora je dikaryotická jarní spora druhů hub z řádu Rustaceae (Uredinales), mající acidiální stadium [5].
- Urediniospora je forma sporulace hub z řádu Rustaceae (Uredinales), po vytvoření aeciospor se vyvíjí na myceliu uvnitř uredinie [5] .
- Basidiospore – exogenní spora, je pohlavní sporulace druhů typu Basidiomycota(Basidiomycota), lokalizované na bazidium vyplývající ze sexuálního procesu [5] [6] .
Biologický význam spór
Podle biologického významu se spóry dělí na dva typy:
- rozmnožovací výtrusy – určené k hromadnému rozmnožování;
- klidové spóry – navržené tak, aby odolávaly nepříznivým podmínkám prostředí [3] .
Propagační spory jsou: zoospory, sporangiospory, konidie, askospory, bazidiospory. Vždy se objevují ve velkém množství a během klíčení mohou produkovat nové spory. Typicky jsou rozmnožovací spory haploidní, začínají haploidní regeneraci a opakují ji několikrát během vegetačního období. Klíčí bez období klidu, někdy i před úplným dozráním, a nejsou schopny dlouhodobého skladování [3].
Klidové spory – cysty, oospory, zygospory, teliospory, chlamydiospory. Tvoří se v malých množstvích, často uvnitř substrátu a nemají žádné prostředky k šíření. Vyznačují se schopností udržet si klidový stav po velmi dlouhou dobu, desítky i více let. Bez stavu dormance nejsou takové spory obvykle schopny klíčit. Dormantní spory obsahují velkou zásobu živin. Dormantní spory jsou diploidní (diploidní stadium), někdy dikaryotické (diaryotické stadium) [3].